Stapeliowe
Stapeliowe – orchidee świata sukulentów
Systematyka
Stapeliowe to grupa sukulentycznych rodzajów należąca, w starszym ujęciu, do rodziny Asclepiadaceae (trojeściowate), a w nowszym systemie do plemienia Ceropegieae podrodziny Asclepiadoideae w rodzinie Apocynaceae (toinowate). Do rodziny Asclepiadaceae należą, obok typowych sukulentów pędowych, także słabsze lub silniejsze kserofity. Liczne są też gatunki tworzące bulwiaste organy spichrzowe – z kolekcjonerskiego punktu widzenia mniej lub bardziej naciągany „kaudeks”. Wszystkie Asclepiadaceae traktowane łącznie jako „sukulenty w bardzo szerokim rozumieniu” stanowią około 900 gatunków, zgrupowanych w 61 rodzajach w ramach rodziny Aclepiadacea liczącej w sumie około 3400 gatunków i 240 rodzajów. Wyraźnie sukulentycznych stapeliowych jest około 330 gatunków.Przykładem interesujących „para-sukulentów” z rodziny Asclepiadaceae, którymi nie będę się szerzej zajmować w tym artykule, są Hoya (200-300 gatunków) i Ceropegia – 180 gatunków. Zwrócę tylko uwagę, że Ceropegia ma bardzo ciekawą, zróżnicowaną morfologię pędów. Bulwiaste organy spichrzowe posiadają Fockea, Cibirhiza, Brachystelma. Wiele gatunków z tej grupy to liany.
We florze Polski stapeliowe nie są reprezentowane. Występują w Azji zachodniej po subkontynent indyjski, w Afryce i południowej Hiszpanii. Ich centrum różnorodności, z największą liczbą gatunków, to Afryka południowa i Wschodnia oraz Bliski Wschód.
Stapeliowe to sukulenty pędowe z silnie zredukowanymi liśćmi. Te są czasem widoczne w szczątkowej postaci na szczycie młodych pędów. Normalne, choć krótkotrwałe, liście posiada jedynie Frerea indica.
Lista sukulentycznych rodzajów Asclepiadaceae, w nawiasie podana orientacyjna liczba gatunków: Baynesia lophophora, Brachystelma (120), Caralluma s.l. (70 gatunków, w niektórych ujęciach dzielone na 17 rodzajów np. Angoluma, Cryptoluma, Sanguilluma, itd.), Ceropiegia (180), Duvalia (18), Duvaliandra dioscoridis, Echidnopsis (32), Edithocolea grandis, Hoodia (14), Huernia (65), Huerniopsis (2), Lavrania haagnerae, Larryleachia (5), Notechinopsis (2), Ophionella (2), Orbea (52), Orbeanthus (2), Pectinaria (3), Piaranthus (8), Pseudolithos (5), Quaqua (30), Rhytidocaulon (10), Stapelia (50), Stapelianthus (7), Stapeliopsis (6), Tavaresia (3), Tridentea (8), Tromotriche (11), White-sloanea crassa.
Dlaczego orchidee świata sukulentów
Stapeliowe swoje miano orchidee świata sukulentów zawdzięczają niesamowitym kwiatom. Kwiaty wszystkich stapeliowych są oparte na tym samym prostym planie. W dodatku jest to symetria promienista, pięciokrotna. Poszczególne elementy okwiatu są modyfikacją gwiazdy pięcioramiennej. Orchidee mają kwiaty o symetrii dwubocznej, grzbieciste. Wydawało by się że taki układ stwarza większe możliwości estetycznego zadziwienia niż symetria promienista. Byłem więc zaskoczony jak, wydawało by się banalna pięciokrotna symetria promienista stapeliowych, może być modelowana w zachwycające konstrukcje.Nauka, o ile taka już istnieje, gdzieś na styku psychologii sztuki, neurofizjologii i psychologii ewolucyjnej, zapewne znajdzie odpowiedzi na pytanie dlaczego symetryczne, promieniste konstrukcje geometryczne (vide mandala, symbole solarne) w szczególny sposób rezonują z naszymi estetycznymi (i nie tylko estetycznymi) emocjami.
Zmienność dotyczy nie tylko kształtu, co mogło by być na dłuższą metę nudne, ale i barw okwiatu i faktury jego powierzchni. Z tego samego geometrycznego wzoru tworzone są nowe jakości estetyczne – wiele gatunków tworzy olbrzymią różnorodność form stanowiskowych różniących się właśnie ubarwieniem i fakturą kwiatów.
Ubarwienie kwiatu jest często pręgowane, marmurkowe – rzecz rzadka u innych roślin. Często występują barwy głębokiej purpury i fioletu – w szczególny sposób oddziałujące na nasz zmysł estetyczny i emocje.
Faktura części okwiatu bywa gładka i lśniąca – wtedy kwiaty wyglądają jakby były wykonane z wosku lub sztucznego tworzywa o głębokiej barwie przenikającej w głąb płatków. Częściej powierzchnia płatków jest pokryta brodawkami, wyrostkami, często jest też owłosiona. Włoski mogą też być krótsze, szczeciniaste, czasem zaopatrzone w buławkowate rozszerzenia na szczycie, wtedy jako tzw. włoski wibrujące (ang. vibratile hairs) poruszają się przy najlżejszym podmuchu powietrza. Z kolei dłuższe i cieńsze włoski tworzą fantastyczne puszyste kwiaty.
[fotki pokazujące zmienność kwiatów stapeliowych] W tym miejscu mogę pokazać tylko niektóre kwiaty stapeliowych. Znacznie więcej możesz obejrzeć na stronie mojej kolekcji www.bio-forum.pl/kaktusy/
Inną analogią z orchideami jest to, że oszołamiającej różnorodności kwiatów towarzyszy względne niewielkie zróżnicowanie morfologii pędów. Te zwykle są nieciekawe i wśród gatunków z jednego rodzaju zwykle niezróżnicowane. Zresztą nawet pomiędzy rodzajami brak często wyraźniejszych różnic.
Podobnie jak u orchidei kwiaty stapeliowych utrzymują się stosunkowo długo. O ile nie dojdzie do udanego zapylenia (co w warunkach kolekcji raczej się nie zdąża) to utrzymują się w zależności od gatunku, od kilku dni do ponad tygodnia. Roślina tworzy zwykle dużo kwiatów w okresie kwitnienia.
Inną analogią do orchidei jest to, że wymagają one specjalnego traktowania w uprawie. Zdecydowana większość stapeliowych jest trudna lub bardzo trudna w uprawie. W przeciwieństwie do stosunkowo dobrze poznanych potrzeb uprawowych poszczególnych grup orchidei – nadal brakuje pewnego „systemu uprawy na stapeliowe”. Szerzej o tym w części poświęconej kolekcjonowaniu stapeliowych.
Budowa kwiatu
Osobliwości budowy kwiatu stapeliowych są wynikiem krańcowego przystosowania do zapylania przez owady, głównie z rzędu muchówek (Diptera). Okwiat wytworzył na drodze ewolucji niezwykłe przystosowania do wabienia określonych gatunków muchówek, różnych dla poszczególnych gatunków stapeliowych i do przeniesienia pyłku w ściśle określony sposób w jedno precyzyjnie określone miejsce.Przy opisie budowy kwiatu podaję także terminy angielskie i niemieckie. Ułatwi to studiowanie opisów gatunków w tych językach, zwłaszcza że różni autorzy stosują nieraz odmienną terminologię. Jej przyswojenie będzie łatwiejsze jeśli czytelnik zrozumie funkcję poszczególnych elementów kwiatu, zwłaszcza tych bezpośrednio zaangażowanych w proces zapylenia.
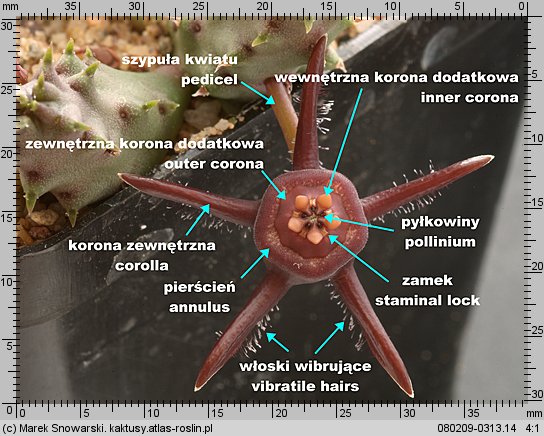
Typowe kwiaty składają się z szypuły (ang. pedicel, niem. Pedicel, Blütenstiel) na której osadzone są zwykle nieciekawe i często niepozorne, zielone działki kielicha (ang. sepals, niem. Sepalen, Kelchblätter), które okrywają, dla odmiany, barwne i atrakcyjne płatki korony. Idąc ku centrum kwiatu znajduje się pręcikowie czyli zespół pręcików (każdy z nitką na której osadzona jest para pylników z osypującym się pyłkiem) i słupkowie ze słupkiem lub słupkami, na szczycie słupka znajduje się znamię słupka, miejsce gdzie przy zapylaniu ma spocząć pyłek. Wewnątrz słupka znajdują się zalążnie z zalążkami. Po udanym zapyleniu i zapłodnieniu z zalążków i innych części kwiatu powstaną nasiona i owoc.
 |
| Huernia hystrix IB8670, widok spodniej strony kwiata |
 |
| Duvalia reclinata Wooldridge IB13768, pędy i kwiat |
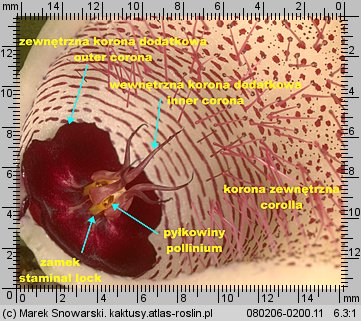
Jej elementy dzielimy na koronę zewnętrzną i koronę wewnętrzną (koronę dodatkową). Korona zewnętrzna (ang. corolla, niem. Kronröhre, Blüte) jest tym co w potocznym rozumieniu kojarzy się z kwiatem. Płatki korony (ang. corolla lobes, niem. Kronzipfel, Blütenzipfel) są duże i w zależności od gatunku różnią się kształtem, barwą, fakturą powierzchni. U niektórych rodzajów (Duvalia, Huernia, Orbea, Orbeanthus) w centrum korony zewnętrznej występuje pierścieniowate zgrubienie (ang. annulus).
 |
| Piaranthus sp. Baden Baden e IB14636, korona dodatkowa |
 |
| Huernia barbata DMC 11539, wnętrze kwiatu po usunięciu części korony zewnętrznej |
 |
| Orbeanthus hardyi, korona dodatkowa |
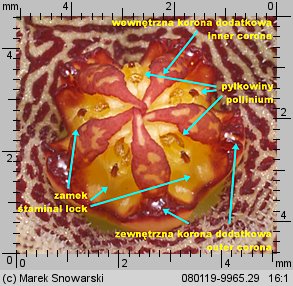
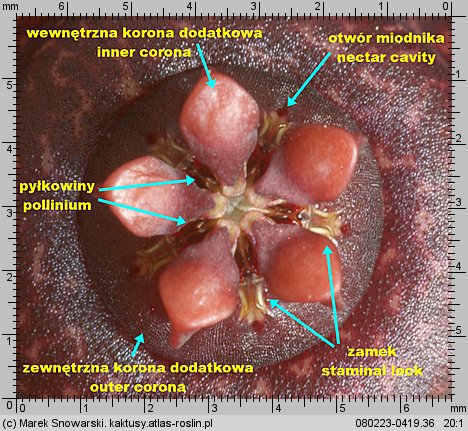
Korona dodatkowa (korona wewnętrzna) (ang. corona, niem. Nebenkrone) jest, z racji niewielkich rozmiarów, stosunkowo niepozorna – jednak jej budowa, kształt i proporcje jej elementów, mają zasadnicze znaczenie w podziale stapeliowych na rodzaje.
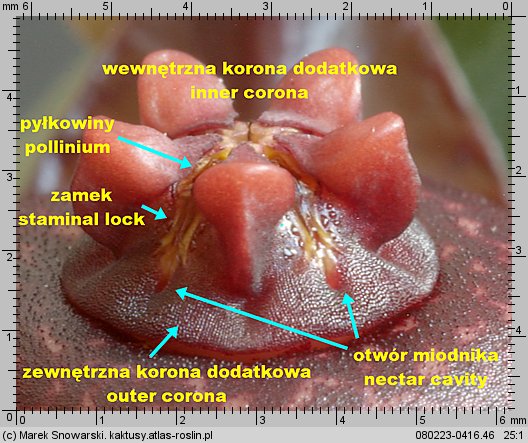
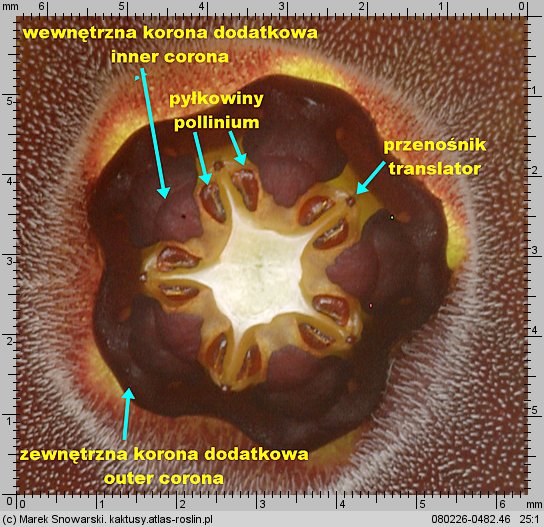
Korona wewnętrzna dzieli się na dwie części – zewnętrzną koronę dodatkową (ang. outer corona, niem. interstaminale Nebenkrone, außere Nebenkrone) – ta jest położona w dolnej części prętosłupa i wewnętrzną koronę dodatkową (ang. inner corona, niem. staminale Nebenkrone, innere Nebenkrone), ta jest położona w górnej części prętosłupa i często ma wywinięte swoje płatki ponad prętosłupem.
 |
| Duvalia polita IB 11026, korona dodatkowa, widok od góry |
 |
| Duvalia polita IB 11026, korona dodatkowa, widok z boku |
Znamię słupka (ang. anther, niem. Anthere) nie jest położone, tak jakby się wydawało na szczycie prętosłupa, lecz leży pomiędzy płatkami korony wewnętrznej, mniej lub bardziej w głębi korony kwiatowej. Jest pięć znamion słupka, tyle ile płatków korony.
Każde znamię ma formę dwóch stulonych warg, tworzących szczelinę. Wargi w dolnej części są rozchylone, ku górze zbiegają się i są zrośnięte w szczytowej części – spełniają one funkcję prowadnic dla odpowiedniego elementu pyłkowiny i zapewniają jej stabilne umocowanie podczas zapylania. Z uwagi na funkcję znamienia będziemy nazywali je zamkiem. Na określenie zamka w literaturze stosuje się różną terminologię jak: ang. staminal lock, guide rail, anther wings.
Poniżej zamka znajduje się otwór miodnika (ang. nectar cavity) – u niektórych rodzajów np. u Piaranthus przy drażnieniu wydostaje się z niego duża ilość lepkiego nektaru.
 |
| Orbea lutea ssp. vaga, korona dodatkowa, widok z góry |
Dwie pyłkowiny są połączone ze sobą przenośnikiem (ang. translator, carrier, niem. Translator, Klemmkörper und Translatoren) z wyrostkami zaczepiającymi się o odnóże owada. Na jednej z krawędzi pyłkowiny znajduje się wydłużony element – klucz (ang. pollinium key, germinating mouth, anchor margin) który, aby doszło do udanego zapyleniu, musi być wsunięty w zamek
Wewnątrz prętosłupa znajdują się zalążnia z dwoma owocolistkami połączonymi jedynie szypułkami. Po skutecznym zapyleniu korona kwiatowa zasycha, odpada, a na szypule kwiatu rozwijają się dwa silnie wydłużone owoce, suche typu mieszek, pękające jednym podłużnym szwem. Swoim ułożeniem owoce przypominają rogi, są stosunkowo duże i długie w proporcji do rozmiaru pędów, osiągają do kilkunastu centymetrów długości. Nasiona są płaskie, obłonione, liczne (kilkadziesiąt), stosunkowo duże i zaopatrzone w puch lotny. Są przystosowane do rozprzestrzeniania przez wiatr. Puch lotny nasion stosunkowo łatwo odpada i nasiono osiada wtedy na podłożu.
Zapylanie
Kwiaty stapeliowych są zapylane przez owady, głownie przez muchówki (Diptera). [mikrofotka pyłkowin i aparatu przenoszącego]W procesie udanego zapylaniu owad zaczepia nogą o aparat przenoszący z przymocowanymi do niego parzystymi pyłkowinami. Odrywa je od prętosłupa i odlatuje do kwiata na innej roślinie tego samego gatunku.
Tam, przemieszczając się w koronie kwiatu, może zaczepić wleczonym na swoim odnóżu, kluczem pyłkowiny o dolną część zamka, szarpiąc i próbując wyzwolić uwięzioną w aparacie przenoszącym nogę może doprowadzić do wsunięcia zamka do klucza do oporu a następnie przy kolejnym szarpnięciu oderwać pyłkowinę od aparatu przenoszącego. Owad może teraz odlecieć z drugą pyłkowiną.
Pyłkowina wsunięta kluczem w zamek pęcznieje w przeciągu około doby. Ziarna pyłka kiełkują – wyrastają z nich łagiewki. Poprzez zamek wrastają one w prętosłup a następnie w zalążnię i przeniesiony w nich materiał genetyczny ojca zespala się z komórkami jajowymi w zalążni. Dochodzi do zapłodnienia. Następnie rozwija się owoc i umieszczone w nim nasiona z zarodkami.
Stapeliowe są w zdecydowanej większości obcopylne. W niewielkim procencie przypadków możliwe jest samozapylenie roślin obcopylnych. W warunkach kolekcji stosunkowo łatwo dochodzi do zapyleń krzyżowych – nawet pomiędzy gatunkami z różnych rodzajów. Na przeszkodzie takim zapyleniom stoi w naturze izolacja geograficzna, izolacja spowodowana wabieniem różnych zapylaczy i niekompatybilność pomiędzy rozmiarem lub kształtem klucza na pyłkowinie i zamka prętosłupa.
Ręczne zapylanie
W warunkach kolekcji praktycznie nie ma co liczyć na zapylenie przez owady. Jedynie sporadycznie dochodzi do przypadkowych zapyleń i należy się liczyć z tym, że nasiona będą wynikiem krzyżówki różnych gatunków kwitnących w tym samym czasie.Pozostaje ręczne zapylanie.
Ponieważ w kolekcjonerskiej uprawie stapeliowych dominuje rozmnażanie wegetatywne, przez sadzonki, to zwykle dysponujemy egzemplarzami jednego gatunku, będącymi klonem. A zdecydowana większość stapeliowych jest obcopylna. Na szczęście u niektórych gatunków próby samozapylenia stosunkowo często kończą się sukcesem.
Aby doszło do skutecznego zapylenia potrzebne jest precyzyjne wsunięcie klucza na pyłkowinie w zamek znamienia słupka. Powtórzę jeszcze raz – precyzja jest konieczna. Podejmowane przeze mnie początkowo, próby naśladowania owada – przez merdanie pyłkowiną w okolicy znamienia, nigdy nie zakończyły się sukcesem. Po bliższym przyjrzeniu się sprawie stało się jasne dlaczego. Szansa na wsunięcie klucza do zamka przy chaotycznych ruchach są mniej więcej takie jak stanięcie monety na jej krawędzi przy rzucaniu „orzeł czy reszka”.
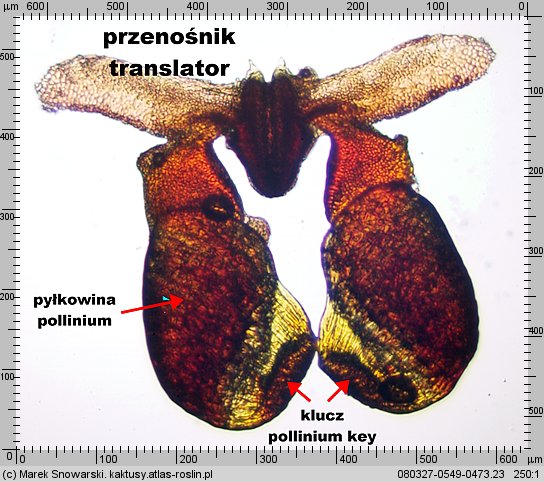 |
| Duvalia polita IB 11069, pyłkowiny z aparatem przenoszącym, widok w mikroskopie w świetle przechodzącym |
Rozmiary zamka i klucza są poniżej 1 mm, zwykle klucz ma długość ledwie 0.2-0.3 mm. Dodatkowo pyłkowiny z przenośnikiem są dość niewdzięcznym obiektem do manipulacji – całość jest dość wiotka i niestabilna. a wyrostki na przenośniku zgodnie ze swoją funkcją czepiają się. Dodatkowo manipulacje utrudnia, zwłaszcza w rodzaju Piaranthus, nektar obficie wydzielany przy dotknięciu zamka. Klucz jest wsuwany w prowadnice zamka „od dołu”, w miejscu do którego dostęp bywa utrudniony – wyjątkiem jest rodzaj Duvalia gdzie całość jest „na wierzchu”. Duvalia jest przez to wdzięcznym obiektem dla ćwiczeń pozwalających nabrać wprawy w ręcznym zapylaniu.
Nie jest możliwe ręczne zapylanie przy użyciu nieuzbrojonego oka. Lupa też się nie zda na nic – za słabe powiększenie i poważna przeszkoda na drodze rąk z narzędziami.
Odpowiednim przyrządem jest mikroskop stereoskopowy (tzw. preparacyjny). Można na nim pracować z powiększeniami rzędu x10 do x50 przy zachowanym stereoskopowym widzeniu (poczucia głębi obrazu).
Oprócz mikroskopu potrzebne są narzędzia do manipulacji pyłkowiną. Chyba najlepiej byłoby sięgnąć do arsenału mikrochirurga. Z braku laku wyprodukowałem proste i skuteczne narzędzia. Do manipulacji użyte są cienkie druciki osadzone na trzonku. Jako drucików użyłem tych które podpierają żarnik żarówki – nie tych drucików na początku i końcu żarnika, są one zdecydowanie zbyt grube, lecz drucika podpierającego żarnik w środkowej części. Bywa on jeszcze zbyt gruby, przez co niewygodny, przy niektórych manipulacjach, tak że warto poszukać innej alternatywy. Jako oprawki użyłem kapilar szklanych do pobierania krwi. Zatopiłem drucik w kapilarze w płomieniu kuchenki gazowej. Oprawka może być użyta dowolna – ważne aby dało się w niej osadzić nieruchomo drucik i aby można ją było w miarę wygodnie trzymać.
Drucik zaginamy stosownie do potrzeb – zwykle przydatne jest zagięcie końcówki w kształcie litery L lub słabsze. Wiele manipulacji można wykonać jednym drucikiem ale łatwiejsze jest to, gdy można też manipulować drucikiem trzymanym w drugiej ręce.
Parę słów o technice zapylania. Zamki są umieszczone z boku prętosłupa – dlatego wygodniej jest, gdy tak umieścimy zapylany kwiat pod mikroskopem, tak aby kwiat był przechylony mniej lub bardziej na bok. U Duvalia pole operacyjne jest całkowicie na wierzchu. U Piaranthus dostęp też jest stosunkowo dobry.
W przypadkach, gdy zamki są umieszczone w głębi korony kwiatowej tak, że są niedostępne, można usunąć koronę zewnętrzną, całą lub niepożądany fragment, przez odcięcie żyletką. Nie zaburza to w najmniejszym stopniu zapylenia a daje nam dostęp do pola operacyjnego. W rodzaju Huernia i Tavaresia, proste odcięcie połowy korony zewnętrznej, w pobliżu jej nasady, daje bardzo dobry dostęp dla manipulacji. Są też trudne rodzaje – np. Orbeanthus, gdzie płatki i wyrostki korony wewnętrznej i niskie umieszczenie zamka dodatkowo utrudnia dotarcie do celu. Można je usuwać przez wyłamanie. Wydzielany obficie sok trzeba wtedy usunąć rogiem chusteczki.
Po przygotowaniu pola operacyjnego przystępujemy do właściwego zapylania.
Po pierwsze musimy pobrać pyłkowiny. Znajdują się one ponad znamieniem słupka. Przyczepione pośrodku przenośnika do prętosłupa. pyłkowiny są złożone bokami ku sobie i przylegają krawędzią do prętosłupa. Wystarczy nacisnąć, przy pomocy naszego drucika, w miejscu przyrośnięcia przenośnika a cały układ pyłkowin odłączy się. Jest on na tyle mały, że będzie dzięki siłom przylegania (a może lepkości) trzymać się drucika.
Po drugie, musimy manipulując drucikiem obrócić pyłkowiny z przenośnikiem. o 180 stopni. Tak, aby były ułożone odwrotnie (tj. przenośnikiem do dołu) niż to było przed odłączeniem od prętosłupa. Takie położenie będzie potrzebne przy wsuwaniu klucza pyłkowiny w zamek znamienia słupka. Pyłkowiny obracamy pomagając sobie drugim drucikiem-manipulatorem. Można też obrót wykonać przy pomocy jednego drucika, opierając pyłkowinę o część kwiatu, choć jest to mniej wygodne.
Po trzecie należy przenieść właściwie ustawioną pyłkowinę, kluczem skierowaną ku zamkowi ku dolnej części zamka i tam wsunąć ją, od dołu do góry, w zamek. Wsuwanie najłatwiej wykonać popychając od spodu pyłkowinę lub, gdy już w miarę pewnie siedzi w prowadnicy zamka, pociągając za przenośnik. Gdy klucz jest wsunięty w zamek, napotkamy pewien opór, który zwykle powoduje rozerwanie przenośnika i pozostajemy z jedną pyłkowiną umieszczoną kluczem w zamku i drugą na druciku. Można ją użyć do włożenia w kolejny zamek. Zamków mamy pięć. Wystarczy umieszczenie pyłkowiny w jednym aby było możliwe zapylenie kwiatu.
Jak widać operacje te wymagają dużej precyzji, drobnych ruchów i początkowe trudności i niepowodzenia mogą budzić frustrację. Z praktyki zapewniam, że jest z tym jak z wkładaniem nitki w oczko igły. Z początku wydaje się niemożliwe. Ale po kilku razach przychodzi w miarę łatwo i operacja zabiera nie więcej jak 3 minuty czasu.
Objawem skutecznego zapylenia jest więdnięcie korony. Mniej więcej po upływie doby od zapylenia. Przy czym szypułka kwiatu, działki kielicha i zalążnia pozostają jędrne, a nawet nieco grubieją. W tym stanie owoc, zanim zacznie wzrost może pozostawać tygodniami, miesiącami, w skrajnym wypadku latami. Trudno przewidzieć jak się zachowa. Im dłużej trwa oczekiwanie tym większa szansa, że pęd na którym wyrósł zamrze z takiego lub innego powodu i nie uzyskamy nasion.
Ale czasem udaje się uzyskać nasiona.
Pędy
Stapeliowe są sukulentami pędowymi. Mają zgrubiałe pędy, z miękiszem wodnym i silnie zredukowane liście – zwykle nie ma po nich śladu lub są jedynie słabo zaznaczone, jako łuski na najmłodszych przyrostach. Pędy mają zwykle niewielką liczbę prosto ułożonych żeber. Ich grzbiety są niemal nagie, pokryte wyrostkami, w rodzaju słabych cierni. Rzadziej zdarza się, że ciernie są stosunkowo pokaźne, żebra liczne i ułożone skrętolegle lub z pozoru nieregularnie.Stapeliowe bronią się przed amatorami ich tkanek gorzkim smakiem (ale raczej nie są silnie trujące) i wydzielaniem obfitego soku mlecznego (wbrew nazwie bezbarwnego), który zasycha, tworząc skorupę w miejscach skaleczeń.
To co ma wielkie znaczenie przy uprawie stapeliowych. to cykl życiowy pędów. W przypadku większości kaktusów (Cactaceae) wzrost pędów jest teoretycznie wieczny – umierają one nie samoistnie, lecz wskutek działalności patogenów lub złych warunków wegetacji. U stapeliowych regułą jest programowana śmierć pędów. Z nielicznymi wyjątkami, pędy żyją od kilku miesięcy do kilku lat, po czym nieuchronnie zamierają – żółkną i wysychają lub, gdy warunki są niekorzystne, to gniją, czasem prowadząc do śmierci całej rośliny. Czasami zdolność do naturalnego zamierania pędów jest przydatna. W ten sposób roślina może odrzucać porażone chorobą pędy – wtedy potrafi utworzyć się sucha tkanka odcinająca na granicy pędu chorego i zdrowego – potrafią to rośliny w dobrej kondycji. W ten sposób roślina może też pozbywać się zbyt dużej powierzchni transpiracji w przypadku gdy brak wody.
Istnieją też nieliczne stapeliowe o teoretycznie wiecznym wzroście pędów – np. rodzaj Hoodia.
Nowe pędy wyrastają zwykle u podstawy młodych pędów starszego pokolenia. Tworząc w ten sposób kroczący łańcuch pędów. Jeśli pędy są wydłużone i wzniesione powstaje wtedy forma krzaczasta, np. u licznych Stapelia. Jeśli pędy są leżące i krótkie, to tworzy się forma płożąca np. u Piaranthus. Część gatunków, np. w rodzaju Huernia ma pędy silnie wydłużone, wznoszące się po podporach lub zwieszające, można powiedzieć, że o charakterze lian.
Kwiaty tworzą się zawsze na najmłodszych pędach. Często w pobliżu podstawy pędów (Stapelia, Huernia), rzadziej w pobliżu szczytu pędów (Piaranthus, Hoodia).
[fotki – typy pędów = Tavaresia, Piaranthus, Huernia, Hoodia, Rhytidocaulon, ]
Kolejny artykuł opowiada o specyfice kolekcjonowania i uprawy stapeliowych.
wersja publikacji 15.12.2011.dvdi · ostatnio zmieniana/last modified 28.10.2008 · została utworzona/was created 28.10.2008
kod html: <a href="http://kaktusy.atlas-roslin.pl/o-stapeliowych.htm">Stapeliowe - kaktusy.atlas-roslin.pl</a>
BBCode na wielu forach: [url=http://kaktusy.atlas-roslin.pl/o-stapeliowych.htm]Stapeliowe - kaktusy.atlas-roslin.pl[/url]